
Поділ клітини: що лежить у витоках цього процесу

Стаття написана Павлом Чайкою, головним редактором журналу «Пізнавайка». З 2013 року з моменту заснування журналу Павло Чайка присвятив себе популяризації науки в Україні та світі. Основна мета як журналу, так і цієї статті – пояснити складні наукові теми простою та доступною мовою.

Як клітини навчилися ділитися? Як? – може здивуватися читач — невже міг бути час, коли клітини не вміли ділитися? Адже це банальна істина, що все живе побудовано з клітин, а клітини розмножуються шляхом ділення. Все це майже в точності так, але все-таки тільки майже: по-перше, хоча всі клітини діляться, але роблять це з різним ступенем досконалості, а по-друге, з клітин побудовано не абсолютно все живе — є ще й віруси.
Всякий вірус складається з нуклеїнової кислоти, компактною спіраллю покладеною в його серцевині, і білкової оболонки. У ньому немає ні набору ферментів, які б робили ці його речовини, ні системи транспорту, яка здійснювала обмін із зовнішнім середовищем, — словом, всього того, що робить клітину клітиною, немає в ньому, зрозуміло, і механізму розмноження. Вірус — не клітина, і щоб розмножитися, повинен потрапити в клітину, де він розмножується без всякого поділу. Поділ клітини — ланцюг неймовірно складних процесів, і саме народження життя стало б набагато зрозуміліше, якби не треба було уявляти перший організм як такий, що вже ділиться.
Може бути, перший організм розмножувався як вірус? Ні, таке припущення не проходить, так як вірус — не організм, він може розмножуватися тільки з допомогою стороннього організму, а його-то при народженні першого організму і не було.
Зате вірус корисний при розумінні народження життя в іншому: він демонструє нам, наскільки по-різному може бути влаштовано розмноження. Якби не віруси, ми напевно вважали б непорушною аксіомою той факт, що єдиним носієм спадкової інформації є двоспіральна ДНК і що всяке розмноження починається з її подвоєння. Однак у вірусах інформація зберігається і в інших формах.
Далі, для виготовлення копії інформаційної молекули як раз подвійна спіраль зовсім не обов’язкова, навпаки, копіювання подвійної спіралі — найскладніша з усіх форм копіювання, що вимагає багатьох ферментів. Адже подвійну спіраль треба спочатку розплести, потім зробити копію з кожної половинки (а напрямки запису і зчитування на двох ланцюгах будь-якої ДНК протилежні, так що і копіювати їх треба з різних сторін) і знову закрутити, тепер вже два двоспіральних ланцюга. Поява такої системи відразу — фантастика. А ось зі звичайною одноланцюговою РНК, яка буває в деяких вірусах, копію може робити один-єдиний фермент. Причому, а це не менш важливо, по РНК можна і будувати основні «цеглинки» — клітини-білки, тоді як з ДНК це виконати неможливо.

Тому для початку нам треба відшукати правдоподібний шлях виникнення перших молекул саме рибонуклеїнової кислоти, тоді подальше поясниться вже значно простіше. Спеціалізований механізм зберігання інформації (так би мовити, «блок зовнішньої пам’яті») міг з’явитися і пізніше.
Отже, парадокс в тому, що найпростіші з живих істот — віруси — зовсім не самі ранні форми життя. Сама ж рання з форм, яку можна собі уявити по «заданим» природою умовам, настільки складна, що ніяк не можна уявити її народження з неживої природи разом.
Залишається йти іншим шляхом — шляхом можливої реконструкції патріарха організмів з найпростіших органічних речовин.
Виходить, що перше живе утворення (його прийнято називати «еобіонт», щоб уникнути терміна «організм», з яким ми звикли пов’язувати щось більш складне, зокрема таке, що володіє системою клітинного поділу) найпростіше представити собі у виді: РНК плюс примітивне пристосування для її зчитування: на цій РНК повинна була бути записана якась інформація, корисна для «виживання» і поширення цієї РНК в «первинному бульйоні». Однак від цього еобіонта до самої примітивної клітини ще неозоро далеко: у клітини завжди є оболонка, що відділяє внутрішнє середовище (цитоплазму) від зовнішньої, причому оболонка вміє вибірково пропускати всередину все, що потрібно цитоплазмі для життя, і назовні — все, що потрібно вилучити; в цитоплазмі міститься все, що потрібно для вироблення енергії, синтезу всіх потрібних речовин, а спадкова ДНК містить коди всіх цих речовин і ще — інформацію про те, які синтези і розпади за якими повинні включатися і виключатися.
Між іншим, все це влаштовано так, що в якийсь момент клітина ділиться на дві, і кожна дочірня клітина отримує весь робочий комплект, та ще й замикається власною оболонкою.
Якщо вважати, що предок перших організмів вже вмів якось ділитися, то доведеться визнати, що його «блок пам’яті» містив коди, принаймні, кількох десятків ферментів і структурних білків, які до того ж виготовлялися в потрібному порядку: крім того, доведеться наділити еобіонт і самим механізмом, які керували поділом клітини. Іншими словами, доведеться визнати, що все найцікавіше вже було зроблено, і ми нічого не в силах сказати про те, як це було зроблено.
Отже, важливо вміти уявити собі еволюцію живого, яке ще не вміло ділитися. Виявляється, це не так вже й важко: адже, по суті, життя найбільш вражаюче не тим, що весь час народжуються нові індивідууми (це роблять і сніжинки при снігопаді), а тим, що воно незбагненно стійкіше і напористіше — весь час захоплює нові території і вперто повертається знову туди, звідки було витіснене. При цьому воно безперервно пристосовується саме до себе, так як «нові території» це перш за все організми, які стають їжею, притулком або грунтом для нових організмів.
Життя немислиме (та й не дуже цікаве) у формі єдиного виду організмів, оскільки той не зміг би нічого більше, ніж з’їсти весь свій «первинний бульйон», включаючи небіотичну сировину. Всі організми і нинішні, і вимерлі, жили і живуть остільки, оскільки інші організми «подають їм на стіл» і «прибирають за ними». Невже саме перше живе утворення було настільки швидке, що управлялося за всіх? Чи означає це, що воно було в якомусь сенсі досконаліше всіх наступних організмів? На це можна відповісти і позитивно і негативно: звичайно, воно було зобов’язане саме себе «обслуговувати», але саме тому навряд чи має сенс порівнювати його з подальшими організмами, так як воно взагалі не могло бути організмом.
Всі без винятку організми володіють тою властивістю, що розмножуються набагато швидше, ніж еволюціонують, але саме ця властивість була б абсолютно неприйнятною для їх попередника, так як привела б до того, що рано чи пізно вся доступна йому частина Землі була б заповнена однаковими голодними істотами (щоб їх знищити або переробити їх речовину, потрібні були б якісь організми іншого типу). Еволюція зупинилася б, як зупинилося окислення металів в той момент, коли на поверхні Землі окислювати стало нічого. Еобіонту набагато перспективніше було б почати не з росту своєї чисельності (розмноження), а з росту різноманітності своїх властивостей. Жоден організм не здатний на це, зате саме так веде себе земне життя в цілому — воно з самого початку присутнє в єдиному екземплярі і весь час збільшує свою різноманітність.
Нинішні організми утворюють біосферу, тобто деяку замкнуту сукупність, в якій попит і пропозиція збалансовані. Якщо допустити, що згаданий «предок» виник в одному примірнику, — що все-таки слід визнати меншим дивом, ніж одночасна поява збалансованої сукупності організмів, то його треба порівнювати не з організмом, а зі всією біосферою. Навряд чи, звичайно, в ньому були точно збалансовані всі можливості і потреби, але абсолютно ясно, що він не міг би і «жити на всьому готовому». Еобіонт потребував безлічі різних хімічних компонентів — нуклеотид, амінокислот, джерела хімічної енергії і т. п., і якщо б він виключно споживав, нічого не виробляючи, він скоро зіткнувся б з нестачею одного за іншим всіх цих речовин: а оскільки можливості його були дуже малі (мала генетична інформація), то неправдоподібно, щоб він міг раптом відразу навчитися справлятися з усією цією лавиною проблем.
Наскільки все виявляється простіше, якщо ця предтеча життя не була клітиною – мікроскопічною крапелькою з набором всього необхідного всередині і з оболонкою зовні; набагато простіше все зрозуміти, якщо слідом за відомим німецьким хіміком М. Эйгеном вважати еобіонт просто деякою системою хімічних реакцій; ця система не була відділена просторово, зате була включена в загальну систему тогочасної геохімії. Наприклад, їй потрібні амінокислоти, а їх, як ми знаємо, в масі поставляє неорганічна природа — вулкани, електричні розряди і т. д.; вона ж тобто природа, і вилучає амінокислоти, включаючи їх в наступні хімічні реакції, розкладаючи сонячним ультрафіолетом, і т. д.
Можна назвати першим еобіонтом ту систему реакцій, яка призвела до первинного накопичення амінокислотних ланцюжків, прообразів білків. Такий «еобіонт-1» був, звичайно ж, включений в систему перетворення речовин Землі, так як брав з неї амінокислоти та їй же їх повертав (хоча б за рахунок ультрафіолетового розпаду). Далі, можна називати «еобіонтом-2» систему реакцій, в яких накопичувалися перші нуклеотидні ланцюжки, прообраз РНК.
Не пов’язуючи ці системи ні з яким районом, а тим більше з крапельками, ми не зустрінемо і труднощів у розумінні того, як вони поєднувалися в просторі, – вони могли переплітатися всюди, хоч по всьому Світовому океану. Як вперше РНК і потрібний їй білок виявилися один одному корисні, ми не знаємо, але можемо досить впевнено припускати, що відповідні хімічні системи були до того моменту вже пов’язані з іншими системами геохімічних перетворень. Не повинно було порушуватися це сполучення і згодом. Ще не розділений на окремі об’єкти, а тому і такий, що не має потреби розмножуватися, еобіонт розростався і завойовував доступний йому простір, тобто в деякому сенсі жив.
Цю концепцію — еволюцію живого шляхом поетапного включення реакцій, що відбуваються до цього поза рамками життя, — її автор, американський біохімік Н. Горовіц, назвав «ретроградною еволюцією», тобто еволюцією, що рухається назад. Термін прижився в науці, хоча фактично він не дуже вдалий: якщо що і відступало, то не життя, а неорганічна хімія Землі, а органічна еволюція весь час наступала.
Реконструкція першої живої істоти, якщо її вести, враховуючи всі умови, які повинно було б дотримуватися життя при своєму виникненні, «видає» нам в результаті якусь неподільну, але таку що еволюціонує напів-істоту напів хімічний самопідтримуючий процес, організм-біосферу.
Роз’єднавши поняття життя, розмноження і клітинного поділу, ми позбулися відразу від декількох труднощів, так як походження життя перестало здаватися фантастичним збігом щасливих випадковостей. Точніше, воно здається тепер не більше (але і не менше!) фантастичним, ніж накопичення, за законами геохімії, певних копалин в певних місцях Землі.
Реєстр переваг такого підходу можна продовжувати і далі: так, всі гіпотези походження життя зустрічаються зі специфічною проблемою, коли хочуть пояснити, як виникла клітинна оболонка. Найчастіше автори пишуть, що оболонка виникла з речовин «первинного бульйону», сама плавала в ньому, немов мильна бульбашка – в повітрі, і виявилася вигідною для еобіонта, що випадково потрапив всередину неї. Це, звичайно, чудово, але не слід забувати, що в цій бульбашці йому і кінець, оскільки він, не будучи скоординований з синтезом оболонки хімічно, не зможе ні рости всередині неї, ні вибратися з неї, ні керувати нею, а значить — і своїм власним діленням.
Щоб уникнути глухого кута, автори цієї гіпотези придумали припущення, що бульбашки, які полонили еобіонтів, могли ділитися під ударами хвиль прибою. Це справді дотепно (перша істота, в разі достатньої спритності, може при цьому втекти зі своєї в’язниці, а автор гіпотези — розлучитися з концепцією «корисної» оболонки), але для еволюції життя потрібна все-таки оболонка, синтезована самим еобіонтом, тобто за законами, записаними у його РНК. Ніякий феномен, випадково виявлений нами в експериментальному бульйоні, не можна прикласти до концепції біологічної еволюції, поки не зрозуміле його хімічне сполучення з усією системою.
Мабуть, первинна оболонка могла виникнути тільки одним шляхом: «предок» збирав її з зовнішнього матеріалу, включаючи в свою систему нові реакції, які до цього проходили в рамках геохімії, і розбирав її, віддаючи цей матеріал назад в те ж геохімічне лоно. Пізніше він, природно, включив в свою систему і виготовлення згаданого матеріалу. Однак важливіше та обставина, що еобіонт, не будучи ізольований просторово, міг, замість повернення речовини в геохімічну систему, передавати його від однієї своєї частини до іншої. Наприклад, він міг використовувати матеріал відслужилої мембрани як джерело енергії, тобто, як би частково з’їдав сам себе — пізніше ці функції відійшли до різних організмів. Іншими словами, коли еобіонту настав час розділитися на різні організми, серед них були вже прообрази і рослин, і рослиноїдних тварин, хижих, і гнильних бактерій — інакше, мабуть, бути не могло. Родоначальник життя, повторюємо, був не першим організмом, а першою біосферою.
Виходить, що живе не відразу стало клітиною, а тому і не відразу постало перед проблемою поділу. До цього воно могло розростатися, не ділячись на організми, і придбати ряд функцій, корисних при клітинному розподілі, ще не обтяжуючи себе механізмом поділу. Зокрема, до клітинного поділу треба було навчитися розмножувати генетичний матеріал — спершу у вигляді РНК, а потім ДНК. Навіщо взагалі знадобилася ДНК? Відповідь (зрозуміло, гіпотетична, як і все, про що ми тут говоримо) дозволяє дати та ж концепція балансу первинного життя з тодішньою геохімією: включаючи в склад своєї РНК все новий і новий нуклеотидний матеріал, еобіонт неминуче був і сам включений в ті шляхи геохімічного круговороту, які розкладали нуклеотидні ланцюги, — інакше еобіонту скоро ні з чого було б будувати нові РНК. Для еобіонта такий оборот подій міг стати трагічним, так як поряд з масовою, надмірною РНК підлягала знищенню і включенню в геохімічний баланс унікальна РНК, що кодувала щойно знайдені еобіонтом «винаходи».
Як не мала відмінність ДНК від РНК, цієї відмінності виявляється достатньо, щоб ДНК могла бути згорнута в більш тугу подвійну спіраль, ніж РНК, – в так звану В-форму ДНК. Тим самим ДНК може перебувати як у формі, схожій на РНК (А-форма), і служити для зчитування інформації з допомогою тих же механізмів, які читають РНК, так і згортатися в нечитабельну В-форму; у цій формі вона і могла ставати неїстівною для каталізаторів, призначених для з’їдання РНК.
Як було вже сказано, система подвоєння ДНК дуже складна, і не з неї почалося життя; але все-таки подвоєння повинно було виникнути до поділу еобіонта на клітини. І зараз, в клітинах, ДНК вміє подвоюватися без ділення самої клітини. Тому головне, що повинен був зробити еобіонт, щоб нарешті розділитися на клітини, це навчитися нарощувати клітинну оболонку, розрізати і зшивати її краї і при цьому забезпечувати кожну нову порожнину копією ДНК.

Предок першої клітини міг вирішити цю проблему приблизно так, як вона вирішена у нинішньої бактерії: вона ділиться, немов перетягаючи сама себе тугим поясом. Перетяжка супроводжується зростанням оболонки — приблизно так росте, закриваючи собою отвір, діафрагма фотоапарата. Коли обидві бульбашки готові, вони самі собою «зашиваються» (діафрагма схлопується) і спливають в різні боки.
Що ж стосується розподілу ДНК, то питання вирішено у бактерій геніально просто: вона завжди до початку поділу оболонки вже подвоєна, обидві двоспіральні копії прикріплені до однієї точки «екватора» клітини, і кожна копія просто вивозиться відповідною ділянкою зростаючою в обидві сторони оболонки.
Словом, головна трудність життя, коли йому настав час утворити з себе окремі організми, — їх одяг, а не зміст.
Як відбувається поділ клітин?
Бактеріям просто: у кожної лише одна двоспіральна молекула ДНК, згорнута в кільце, і лише одна оболонка — та, що утворює саму бактерію. В інших клітинах все влаштовано набагато складніше: всередині клітинної оболонки є інша оболонка — ядро, і ДНК розташоване саме в ядрі, причому вона представлена не однією молекулою, а кількома дуже складними конструкціями — хромосомами. Кожна хромосома до початку поділу клітини подвоюється, тобто клітина виготовляє замість неї дві точно таких же хромосоми. Це саме по собі диво (адже хромосома — не просто молекула ДНК, а складне переплетення ДНК і білків) і дуже ефектне зовні: в спочиваючій клітині хромосоми не видно, і перед тим, як подвоюватися, вони ніби заново народжуються всередині ядра; але потім починають змінювати одна одну ще більш ефектні картини.
Ядерна оболонка розчиняється, але хромосоми, опинившись на волі, не біжать хто куди, а як би шикуються в дві шеренги, причому кілька сусідок утворюють завжди дві сестри, що походять від однієї хромосоми. Цікаво, що цей лад розташовується не де попало, а посеред клітини, уздовж майбутнього екватора, за яким почне ділитися клітинна оболонка. Потім шеренги починають чітко розходитися до протилежних полюсів клітини, так що сестри назавжди залишають одна одну. Управляє цим рухом, як в театрі маріонеток, пара пучків найтонших ниточок, що сходяться до полюсів, – веретена поділу. Коли хромосоми покинуть екваторіальну область, клітина починає звужуватися, її як би стягує тугий пояс (це, втім, відбувається тут так само, як у бактеріальної клітини), і вона ділиться на дві. Нарешті в кожній новій клітині навколо хромосом виростає нова ядерна оболонка, і хромосоми знову стають невидимими, як би розчиняються. Такий типовий механізм клітинного поділу – мітоз.
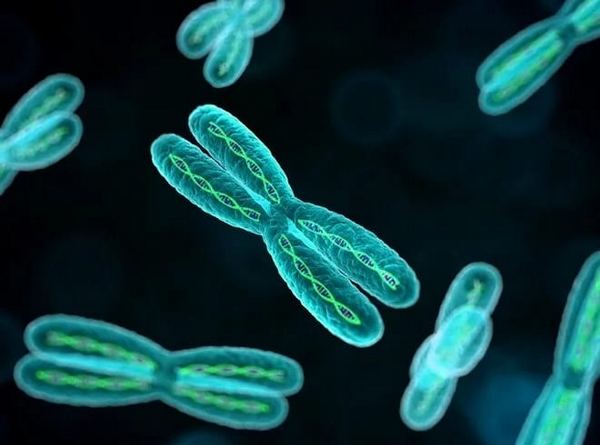
Про поділ клітини зняті фільми, він яскраво і натхненно описаний багатьма, але всякого дослідника, який спостерігає за ним, він вражає заново — маленьке диво, що повторюється з дивовижною точністю з безліччю клітин у кожної з незліченної безлічі організмів.
Сто років тому, коли мітоз був відкритий і детально вивчений, вчені були переконані, що він загалом протікає однаково в усіх організмах — у тварин, рослин і навіть в одноклітинних організмах, наприклад у інфузорій. Все добре, крім одного — було абсолютно незрозуміло, як такий фантастичний механізм міг відбутися. Однак на початку XX століття стало з’ясовуватися, що картини мітозу, типові для більшості багатоклітинних, далеко не так звичайні серед одноклітинних; скоріше можна визнати, що типовий мітоз зовсім не типовий для одноклітинних і що інфузорії, швидше за все, стоять тут осібно.
Перш за все, виявилося, що у багатьох одноклітинних ядерна оболонка не розчиняється в ході поділу, а ділиться навпіл, як і сама клітина, причому веретено поділу розташовується усередині ядра. Народилася гіпотеза, згідно з якою ядро — це бактерія, що живе паразитом всередині клітини, а розчинення ядра в ході типового мітозу — більш пізніше еволюційне придбання. Слів немає, гіпотеза була на ті часи красива, але насторожувало те, що ніхто не міг вказати такої клітини — щоб ядра у неї не було, а хромосоми і мітоз були; не відомо таке і в даний час.
Потім були виявлені одноклітинні (в основному — водорості), у яких мітоз організований ще дивніше: хромосоми розходяться всередині ядра, яке ділиться, не розчиняючись, але веретено поділу дивним чином розташовується поза ядра, пронизуючи своїми нитками ядерну оболонку: або, що вже зовсім незрозуміло, мітоз, в якому хромосоми розходяться взагалі без допомоги веретена. Нарешті, знайшлися і такі одноклітинні, у яких і хромосоми не цілком справжні між мітозами не розчиняються, а видні завжди.
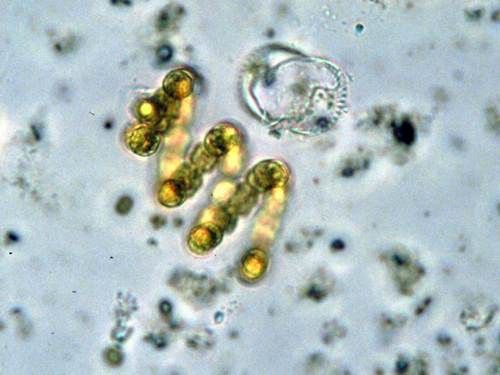
На останньому слід зупинитися докладніше. Найпримітивніші хромосоми (і найпримітивніший поділ) виявлені у одноклітинних водоростей динофлагеллат (що означає «панцирні жгутиконосці») — хромосоми пухкі, схожі на недбало змотані мотки мотузок. Після багаторічних кропітких експериментів і аналізу сотень електронних мікрофотографій вчені прийшли до переконання, що динофлагеллатна хромосома — не що інше, як сукупність безлічі кілець ДНК, кожне з яких схоже на ДНК бактерій. При поділі ці хромосоми розводяться до полюсів поділу ядра не нитками веретена (воно, як таке, взагалі при такому розподілі не утворюється), а як би повзуть за розтягнутою оболонкою. Розповзаються хромосоми не так синхронно, як при типовому мітозі, і до того ж весь процес клітинного поділу займає тут у багато разів більше часу.
Зрозуміло, відразу ж з’явилася гіпотеза клітинної еволюції, згідно з якою панцирні жгутиконосці — найдавніші і найпримітивніші з клітин, що мають ядро. (Про те, як вони могли з’явитися, ми поговоримо нижче.) Наступний етап еволюції – поділ, що спостерігається у водоростей евглен, теж примітивний, але вже з компактними, а не пухкими хромосомами. (Ця гіпотеза стала ще привабливішою, коли в 1974 році з’ясувалося, що при поділі деяких панцирних жгутиконосців рухом хромосоми насправді управляє ядерна оболонка, майже так само, як при поділі бактерії.) Потім повинно було з’явитися веретено, пізніше воно стало виходити з ядра через дірки; нарешті, механізм, який створював ці діри, міг бути використаний і для повного розчинення ядерної оболонки при діленні.
Однак незабаром з’ясувалося, що в такому ряду не знаходиться місця для багатьох форм мітозу; крім того, в межах однієї і тієї ж групи — динофлагеллат — представлені і майже всі інші типи розподілів; крім описаного примітивного мітозу, у них відомі види з нормальним внутрішньоядерним мітозом і навіть з розчиненням ядра; у деяких динофлагеллат і хромосоми компактні. Невже динофлагеллати самі «винайшли» всі ті мітози, які відомі в природі?
Тут, може бути, правильніше буде змінити питання, тобто цікавитися не рядом послідовних організмів з різними мітозами, а прямо — перетворенням самих розглянутих нами структур і функцій: ми, наприклад, не сумніваємося, що наша рука пішла від передньої лапи чотириногого, а та, в свою чергу,— від плавника риби, хоча йдуть нескінченні суперечки про те, що це були за чотириногі та риби. Перебравши різні варіанти, винайдені клітинами для свого поділу, ми переконуємося, що їх ніяк не збудувати в порядок — від простого до складного, щоб він до того ж відповідав іншому — від раннього до пізнього.
Спробуємо підійти до проблеми: чи міг взагалі механізм мітозу, навіть самого примітивного, скластися з тих механізмів, які ми знаємо у без’ядерних клітин? У примітивному мітозі ядро ділиться подібно бактерії, отже, тут міг працювати вже відомий бактеріальний механізм; примітивні хромосоми схожі, як ми бачили, з пучком бактеріальних ДНК, отже, і їх могли поставити бактерії; від них же — бактеріальний принцип роз’єднання хромосом в ході примітивного мітозу.
Веретена поділу тут немає, але навіть при самому примітивному мітозі вдається виявити кілька ниточок; вони тягнуться від одного полюса поділу до іншого і служать, мабуть, направляючими полозами для рухомих хромосом (адже хромосом кілька, і кожна набагато більше, ніж бактеріальна ДНК); вони ж, мабуть, служать для орієнтації поділу ядра клітини. В електронному мікроскопі видно, що кожна ниточка – порожниста всередині, тобто є мікротрубочкою. Мікротрубочок ж у світі бактерій не знайдено.
Залишається спробувати знайти що-небудь схоже на ті компоненти, з яких будується мікротрубочка. У дуже сильний електронний мікроскоп можна побачити, що всяка мікротрубочка складена, немов кабель, з мікрониток, складених, в свою чергу, з білкових кульок. Майже у всіх організмів мікротрубочка складена з 13 мікрониток. Але у примітивних організмів – дріжджів – знайдені мікротрубочки по 7 мікрониток; а ще більш прості і пухкі «кабелі» нещодавно вдалося побачити і у без’ядерних клітин, ціанобактерій (раніше їх називали синьо-зеленими водоростями), які вже давно імовірно вважалися предками рослин.
Тепер у першому наближенні все виходить добре: ми здатні подумки «зібрати» примітивний розподіл ядерних клітин з бактеріальних структур, так що не доводиться припускати таємничої одночасної появи відразу багатьох нових структурних «цеглинок». Більше того, ми вже здатні тим же шляхом «зібрати» і типовий мітоз вищих організмів.
Отже, якщо не «оглядатися» на реальну природну історію, а реконструювати елементарний ряд форм, що ускладнюються, вибираючи їх довільно, виявиться, що ми, власне, знаємо, як клітини навчилися ділитися.
Так вже прийнято у еволюціоністів – вважати, що походження зрозуміле, якщо вказано, хто від кого пішов, хоча фактично в такій вказівці може бути дуже мало розуміння. Що ми можемо відповісти на питання, хто від кого пішов в даному випадку? Досі подумки збираючи механізми мітозу, ми користувалися даними про найрізноманітніші організми: про бактерії, ціанобактерії, про різні типи водоростей, про нижчі гриби і нижчих тварин. Що ж відомо про походження цих організмів? Чи можна скласти такі дані з гіпотезами про походження мітозу?
Тут нічого не виходить: якщо що і походить від ціанобактерій, так тільки водорості багрянки, а ніяк не дріжджі; самі ж багрянки – тупик еволюції. Евглени і динофлагеллати теж далекі один від одного, дріжджі — це взагалі, за сучасними поглядами, не рослини, так що доводиться задати питання: на якій підставі ми використовуємо всі ці дані при демонстрації якоїсь певної еволюції?
Нарешті, якщо мітоз складними шляхами досяг спостережуваної досконалості в царстві рослин, то чому майже точно такий же мітоз ми спостерігаємо і у вищих тварин? Перша відповідь, яка спадає на думку, така: типовий мітоз вищих організмів чимось вигідніший, тому він і представлений у всіх вищих, тобто типовий мітоз сформувався незалежно у рослин і тварин під дією природного відбору.
Однак відповідь не витримала ніякої критики фактами: динофлагеллати з самим примітивним мітозом належать до найбільш поширених на Землі океанічних організмів, тоді як динофлагеллати зі складним мітозом — убогі внутрішньоклітинні паразити: у морському планктоні прекрасно співіснують клітини з усіма типами мітозу, заповнюючи всі тамтешні екологічні ніші (всілякі температури, солоності, умови харчування тощо). Більш того, типовий мітоз характерний для переважної більшості, але все-таки не для всіх високорозвинених організмів — причому тут відбір?
Взята ізольовано, ця проблема видається неприступною, але виявляється, що вона ж встає буквально в будь-якому розділі біології, тобто рішення слід шукати в більш загальному вигляді, безвідносно до еволюції клітинного поділу.
Зокрема, сучасне розмаїття властивостей організмів може давати нам матеріал для уявного відтворення еволюції тієї чи іншої конструкції, не даючи достатнього матеріалу для відповіді на питання, чим ця конструкція вигідна і хто від кого пішов. Відповісти на нього вдається тільки в тому випадку, якщо брати одні ознаки і ігнорувати інші. Донедавна кожен систематик так і вважав, що його завдання — знайти більш вдалий набір ознак, ніж ті, що були знайдені його попередниками, і тим самим вказати більш ймовірного предка даної групи. Проте останнім часом стала з’ясовуватися більш фундаментальна обставина: якщо еволюція більшості органів і структур не йшла по прямій, то нинішнє розмаїття організмів просто не несе достатньої інформації про те, хто від кого походить. Звичайно, це прикро, однак це не завжди так вже заважає нам зрозуміти, як відбулися фундаментальні механізми живого, і зокрема — як клітини навчилися ділитися.
Автор: Ю. Чайковський.