
Нервово-м’язовий синапс: його будова, збудження, принцип роботи

Стаття написана Павлом Чайкою, головним редактором журналу «Пізнавайка». З 2013 року з моменту заснування журналу Павло Чайка присвятив себе популяризації науки в Україні та світі. Основна мета як журналу, так і цієї статті – пояснити складні наукові теми простою та доступною мовою.
Зміст:
Очевидно, що якби всі люди одночасно перестали і розуміти, говорити, то замовкли б заводи і фабрики, встали поїзда, розпалися держави, зникла цивілізація. Але ж будь-яка людина — свого роду держава теж, густо заселена громадянами-клітинами. Кожна з них — індивідуальність, кожна дихає, працює, їсть. Але жити окремо — як це роблять, наприклад, інфузорії — наші клітини вже не можуть. Вони безперервно передають один одному й іншим клітинам свої повідомлення. Але як? За допомогою так званих синаптичних рецепторів.
Визначення
Мабуть, тут потрібно пояснювати кожне слово. «Синапсом» називається контакт між нервовими клітинами або між нервовим закінченням і м’язовим волокном. «Рецепторами» в біології зазвичай називають чутливі утворення ока, вуха, носа. Але тут мова йде про інше значення цього слова. Молекулярні рецептори — білкові молекули, які вловлюють спеціальні речовини-передавачі (їх називають медіаторами), що виділяються з нервових закінчень.
Як й інші клітини, нервові клітини в лабораторних умовах живуть в сольовому розчині, що нагадує за своїм складом морську воду: в ньому багато іонів натрію, хлору і калію мало. Зовнішній розчин відділений клітинною мембраною від внутрішньої частини клітини — протоплазми, в якій, навпаки, багато іонів калію і мало — натрію і хлору. Мембрана нейронів має чудову здатність пропускати через себе одні іони і не пропускати інші. Коли нейрон знаходиться в спокої, його мембрана вибірково проникна для іонів калію. Коли проникність змінюється, і притому певним чином, нейрон генерує нервовий імпульс-сигнал, який він передає іншій клітині по нервовому волокну.
Закінчення нервових волокон підходять до нейронів і м’язових волокон дуже близько, але не впритул. Цей контакт двох клітин і є синапс.
Види
Синапси бувають двох типів: хімічні та електричні. У синапсі між мембранами двох клітин залишається щілина шириною в кілька тисячних часток міліметра. Нервовий імпульс не може через неї «перескочити» і, щоб подолати «перешкоду», нейрон, що передає сигнал, виробляє спеціальну речовину-медіатор, який впливає на мембрану наступної клітини і змінює її проникність для іонів. В результаті наступна клітина починає генерувати нервовий імпульс.
Передача збудження
Механізм дії медіатора був вивчений у серії робіт англійського вченого, лауреата Нобелівської премії Б. Катца і його співробітників, які в 1953 році виявили, що медіатор виділяється з нервових закінчень порціями. Кожна така порція викликає на мембрані «відповідальної» клітини слабку зміну потенціалу, яку зазвичай називають «мініатюрним потенціалом». Пізніше за допомогою електронного мікроскопа, розділивши попередньо синаптичні нервові закінчення на частини, вдалося з’ясувати, що медіатор в нервовому закінченні щільно упакований в маленькі синаптичні бульбашки.
Безліч таких бульбашок плаває всередині закінчення. Коли бульбашка прилипає до зовнішньої мембрани нервового закінчення, вміст такої бульбашки — «квант» збудження — виділяється в синаптичну щілину. Так виникає один мініатюрний потенціал. Кванти виділяються і в спокої, але Катц показав, що нервовий імпульс в тисячі разів збільшує їх середню частоту, не змінюючи величину окремої порції.
Але чому нервовий імпульс прискорює злипання синаптичних бульбашок з зовнішньою мембраною нервового волокна? Спочатку здавалося, що причина очевидна. Нервовий імпульс, приходячи до мембрані, зменшує на ній різницю потенціалів. А ще раніше було відомо, що в сольовому розчині всілякі дії, що зменшують цю різницю потенціалів, збільшують частоту мініатюрних потенціалів. Але були поставлені інші досліди, в яких із зовнішнього розчину видаляли іони кальцію, і тоді ніякого збільшення частоти не спостерігалося, незважаючи на прихід нервового імпульсу. І навпаки, різко збільшити частоту виділення квантів можна без усякого імпульсу — треба лише підняти концентрації в зовнішньому розчині будь-яких непроникаючих іонів або нейтральних молекул, наприклад сахарози.
Ця заплутана ситуація поставила в глухий кут англійських дослідників. Розплутати її вдалося авторам «кальцієвої гіпотези» вченим Е. А. Ліберману і його співробітникам. Вони виходили з простої фізичної ідеї: для злипання бульбашок з зовнішньою мембраною треба зменшити поверхневий електричний заряд мембран — а це роблять іони кальцію, які входять в нервове закінчення з-за того, що приходить нервовий імпульс. Тому-то в розчині, позбавленому кальцію, немає синаптичної передачі.
Оскільки діаметр нервового закінчення зазвичай менше мікрона, ввести туди іони кальцію для прямої перевірки гіпотези не можна. Але дослідники скористалися тим, що кальцій в нервових закінченнях завжди є у великих кількостях всередині мітохондрій, де його тримає електричне поле цих внутрішньоклітинних «електростанцій». Їм це дозволило, викидаючи кальцій з мітохондрій різними агентами, що знімають електричне поле, «ввести» кальцій всередину нервового закінчення. Виявилося, що всі речовини, що знімають електричне поле на мембрані мітохондрій, різко підвищують частоту виділення квантів ацетилхоліну.
Надалі естафета знову перейшла до англійців — їм вдалося поставити прямі досліди на гігантських кальмарів синапсах, в яких товщина нервового волокна досягає міліметра. Так була підтверджена гіпотеза Е. А. Лібермана про роль кальцію в синаптичній передачі. (Проте яким чином розкривається бульбашка, щоб вилити свій вміст в синаптичну щілину, як його мембрана стає частиною поверхневої мембрани нервового волокна — ці питання досі не мають відповіді.).
Після того як медіатор відпрацював, його потрібно прибрати. Для цього існують різні механізми. Один з них — ферменти-руйнівники, молекули, які сидять у синаптичній щілині медіатора і розрізають на шматки. Інший — клітина дбайливо збирає медіатор, щоб використовувати його знову. Вона втягує всередину себе частину мембрани, отриманий пухирець «відшнуровується» та захоплює зовнішній розчин з усім його вмістом. Такий механізм зручний не тільки для клітини, але і для вчених, які його вивчають. Використовують його так: зі звичайного хрону виділяють фермент-руйнівник піроксидазу і вводять його в район синапса. Нервове закінчення заковтує піроксидазу разом з медіатором. Нейрони, що послали свої відростки у місце, куди була введена піроксидаза, тепер легко виявити за спеціальним забарвленням, яке виникає через те, що піроксидаза, працюючи, розщеплює перекис водню і виділяє молекулярний кисень. Так складають карту зв’язків між нейронами і розбирають принцип роботи синапсу.
Будова
Синапс складається з трьох основних елементів:
- Пресинаптичної мембрани, яка покриває розширене нервове закінчення, що представляє собою нейросекреторний апарат. У пресинаптичній частині знаходяться бульбашки та мітохондрії, що забезпечують синтез медіатора. Медіатори депонуються в гранулах (бульбашках).
- Постсинаптичної мембрани – це потовщена частина мембрани клітини, з якою контактує пресинаптична мембрана. Вона має іонні канали і здатна до генерації потенціалу дії. Крім того, на ній розташовані спеціальні білкові структури — рецептори, що сприймають дію медіаторів.
- Синаптичної щілини, вона являє собою простір між пресинаптичною та постсинаптичною мембраною, заповненою рідиною, близькою за складом до плазми крові.
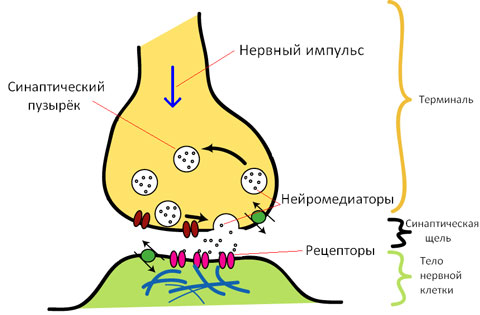
Ось так будова синапса виглядає на малюнку.
Медіатори
Медіаторів зараз відомо багато, і число їх постійно зростає. Зазвичай це порівняно прості хімічні речовини — серотонін, допамін, норадреналін, але медіаторами є багато і окремих амінокислот, а також сполуки, що складаються з декількох амінокислот,— поліпептиди. Також медіатором служить ще і всім відома молекула АТФ. Воістину дивно різноманітна роль цього з’єднання! Воно і основний енергетичний резерв клітини, і служить для синтезу РНК, і бере участь в синтезі білків і в м’язовому скороченні, і використовується для фосфорилювання різних білків і бере участь в іонному транспорті. А тепер ось виявляється, що АТФ — ще й медіатор, тобто виділяється з нервового закінчення, сигналізуючи: прийшов нервовий імпульс.
Важко довести, що така «багатостороння» речовина, як АТФ, є медіатором. Всі необхідні в цьому випадку перевірки надзвичайно ускладнюються. Перш за все, щоб переконатися, що дана речовина є медіатор в даному синапсі, потрібно показати, що воно виділяється з нервових закінчень у відповідь на нервовий імпульс. Однак всюдисуща АТФ може виділятися не тільки з досліджуваних нервових закінчень, але і таких, що знаходяться поруч з чутливими нервовими волокнами і навіть з м’язових волокон!
Інша обов’язкова процедура – розділити синапс на частини і спробувати виявити медіатор в синаптичних бульбашках. Але АТФ є у всіх частинах клітини! Нарешті, третій необхідний доказ – показати дію на постсинаптичну мембрану. Але і тут АТФ може відігравати іншу роль: служити джерелом енергії.
Бернштоку та його співробітникам потрібно було поставити безліч тонких і трудомістких експериментів, щоб отримати серйозні аргументи на користь своєї гіпотези. Серед інших медіаторів особливе місце у ацетилхоліну. Справа не в тому, що він був відкритий першим і на ньому були отримані основні відомості про механізми синаптичної передачі. Просто «медіаторів дідусь» — це поки що єдиний, для якого виділені і добре очищені білкові рецептори — речовини, що безпомилково вловлюють його.
Вченим тут дуже пощастило: природа створила для них електричного ската.

Кілограм тканини його електричного органу містить до ста міліграмів рецепторного білка, що в тисячу разів більше, ніж в м’язах тварин. Очищені рецептори ацетилхоліну з електричного органу ската були вбудовані в штучні біомолекулярні мембрани. Ці мембрани під дією ацетилхоліну міняли проникність для іонів натрію. Експеримент можна грубо трактувати так. Молекула-рецептор містить в собі канал, ворота якого зазвичай закриті. Медіатор змінює форму білка-рецептора відкриває і ворота для іонів – через відкритий канал в клітину йде натрій.
Іонний канал в мембрані – непростий пристрій. У водному розчині іони оточені диполями води. Щоб пройти мембрану, потрібно скинути водну оболонку, але енергетичні витрати на це дуже великі. А канал вміло підміняє собою диполі води. Іон роздягається «без труднощів» і, пройшовши канал, акуратно надягає нову водяну шубу. У результаті іони можуть проходити крізь мембрану за рахунок однієї лише енергії теплового руху.
Штучна система, що складається з мембран з вбудованими білковими рецепторами, була детально описана вченим з Франції Ж. Л. Попо. На жаль, штучна система поки з незрозумілих причин погано моделює нервово-м’язовий синапс: препарати, які на живих об’єктах чітко блокують дію ацетилхоліну, в штучних системах викликають протилежний ефект.
Природа створила багато медіаторів. Але цього їй здалося мало, вона і зробила так, що один і той же медіатор викликає до того ж у різних нейронів різні відповіді.
Француженка Жак Сю Кехое знайшла цілих три типи відповідей на ацетилхолін у морського молюска аплізії. Ця невелика тварина стала класичним об’єктом нейрофізіології. Справа в тому, що для реєстрації відповідей нейронів потрібно, проткнувши мембрану, увійти всередину нейрона мікроелектродом – маленькою скляною піпеткою, заповненою розчином електроліту. Кінчик її не більше мікрона в діаметрі, але біда в тому, що розміри звичайних нейронів не перевищують 30-50 мікрон, і тому для клітини введення електрода — те ж, що для людини удар шпагою. А сучасна постановка досліду вимагає вводити в клітину два і навіть три електрода. Крім того, нервові клітини у більшості тварин заховані в глибині нервової тканини, шукати потрібний нейрон і вводити в нього електрод доводиться наосліп.

Молюск аплізія.
Аплізія ж позбавлена всіх цих недоліків: діаметр її нейронів — сотні мікрон, а нервова система влаштована так, що клітини лежать на поверхні і так чітко видні, що вчені впізнають багато з них в «обличчя»: нейрони володіють характерним забарвленням, розмірами, положенням серед інших клітин, типом активності. Важко описати задоволення, що отримується при роботі з нейроном, який одночасно можна бачити.
Можливо, що коли-небудь аплізії, як і павлівським собакам, поставлять пам’ятник. Якщо вдасться зберегти в ньому натуральні кольори цього молюска, то споруда вийде не тільки вражаючою, але і красивою.
Отже, саме у аплізії Кехое досліджувала реакцію нейронів на ацетилхолін. Всередину нейрона вводила вона два електроди: один для реєстрації, інший для пропускання струму через мембрану. Крім того зовні до клітки підводили спеціальний електрод, заповнений розчином ацетилхоліну. Цей електрод грав роль нервового закінчення: з нього певними порціями можна було випускати позитивно заряджені молекули медіатора. Кехое отримала відповіді на ацетилхолін, які відрізнялися і за величиною, і за тривалістю, і за швидкістю виникнення. Крім того, різні відповіді зникали під дією різних отрут. Вдалося встановити, що різні типи відповідей були пов’язані зі змінами проникності мембрани для різних іонів.
Фізіологи знайшли, що ще більшу різноманітність відповідей викликає інший медіатор – серотонін. Ці роботи були зроблені як на аплізії, так і на нейронах виноградного равлика.
Група нейронів равлика може з часом змінювати свої рецептори серотоніну і, внаслідок цього, характер своєї відповіді на серотонін. А тип відповіді залежить від того, прокинулася равлик після зимової сплячки або ще «дрімає».
Отже, нейрони один з одним пов’язані, нейрони один з одним «говорять». Дослідники спостерігають за їх спілкуванням і намагаються зрозуміти його механізми. Відомо досить багато: як клітини «ворушать губами», як «відкривають рот», як «видають звуки»… Ми знаємо: зараз ця клітина відповідає іншій. Але чи зрозуміємо ми коли-небудь, про що вони говорять?
Автор: С. Мініна.
Рекомендована література та корисні посилання
- Савельев А. В. Методология синаптической самоорганизации и проблема дистальных синапсов нейронов // Журнал проблем эволюции открытых систем. — Казахстан, Алматы, 2006. — Т. 8, № 2. — С. 96—104.
- Экклз Д. К. Физиология синапсов. — М.: Мир, 1966. — 397 с.
- French R.D. Some problems and sources in the foundation of modern physiology in Great Britain // Hist. Sci.. — 1971. — № 10. — С. 28-29.