
Еволюційна теорія та біологічна еволюція

Стаття написана Павлом Чайкою, головним редактором журналу «Пізнавайка». З 2013 року з моменту заснування журналу Павло Чайка присвятив себе популяризації науки в Україні та світі. Основна мета як журналу, так і цієї статті – пояснити складні наукові теми простою та доступною мовою.
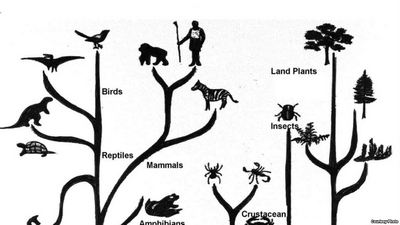
Зрозуміти, як розвивається живе, не можна без розуміння, як воно організовано. Тому почнемо з того, що життя складне. Або, як кажуть нині в науці, це складна самоорганізована система. А самоорганізуються системи — найскладніші, і наука ще не навчилася впевнено в них розбиратися.
Умами біологів поступово опановує думку про те, що «життєва» складність не може бути зведена до будь-якої однієї форми або системи. Ця думка стає одним з наріжних каменів в теоретичному фундаменті біології.
Супроводжується цей процес відмовою від ідеї, яка ще недавно була в числі святая святих біологів: організм, що споконвіку вважався єдино первинною організованою формою життя, втратив право на пріоритет. Зміцнюється переконання, що в живому покриві Землі спочатку є безліч підлеглих один одному систем, що володіють різними ступенями організованості…
Дивно, що ще зовсім недавно безроздільно панував погляд зовсім інший. Порівнюючи його з нині панівним, залишається дивуватися рішучості того, що відбувається в біології “перегляду” її основ. Ще й років тридцять тому цілісною “одиницею” живого визнавалася тільки особина. Надорганізмові системи, біоценози або популяції, скажімо, не вважалися такими, що їх зводили до суми особин, або навіть зовсім заперечували їх існування.
Подібні помилкові тлумачення (вони і нині зустрічаються) витоком своїм мають недостатність знань про складність живого. Досі дуже поширені спроби створювати єдині ієрархії живих систем за типом «драбини», що призводить до відома воєдино різнорідних явищ природи, систем абсолютно різних за ступенем універсальності, як здатних, так і не здатних самостійно існувати й відтворювати собі подібних. Такою однобічністю грішать навіть найвизначніші теоретики. Ось, скажімо, систематизація, запропонована одним з найбільших дарвіністів, Іваном Івановичем Шмальгаузеном: первинна жива частка, клітина, організм, популяція, вид. Ясно, що в цій системі перші три назви «належать» іншому ряду, ніж останні два.
Подібні ієрархії будували багато, але, як правило, немов в тестах, що визначають здібності до логічного мислення, в них завжди можна відшукати зайві ланки, що належать «іншому» ряду.
Насправді життя організоване в кілька ієрархічних рядів зіставлених систем. Притому не всі системи пов’язані між собою саме ієрархічно. Думку про те, що спочатку життя було організоване в кілька форм, висунув І. Вернадський. Він стверджував, що вкорінений погляд на “первородність” організму і вторинність, похідність всіх інших форм життя не відповідає біогеохімічним закономірностям. Процес виникнення життя повинен був відразу втілити його в комплекс складних форм. Первинні особини створювалися природою одночасно з первинними видами, біоценозами і біосферою. Надорганізмові системи, таким чином, настільки ж древні, як і саме життя.
Дивно, але «монополія» організму в біології була настільки стійкою, консерватизм тут виявився настільки наполегливим, що ці ідеї довго залишалися в тіні. Саме тому концепція про одночасне становлення кількох форм організації живого (організмової, популяційно-видової, біоценотичної та біосферної) так повільно завойовує позиції.
Ідеї Вернадського отримали визнання в чомусь завдяки розвитку еволюційної теорії популяційного підходу; з часом одиницею еволюції стала вважатися популяція, а не особина, в біоценології біоценоз був визнаний цілісною системою, що не зводилася до групи індивідів, в теоретичній біології стала популярною загальна теорія систем.
Серед численних форм організації живого важливо розрізняти основні, тобто первинні высокосамостійні, універсальні, стійкі системи, пов’язані між собою ієрархічними і сітчастими відносинами. Саме вони визначають існування і еволюцію життя. Це організмова, популяційно-видова, біоценотична та біосферна форми.
Тут, правда, треба підкреслити одну характерну властивість живого, що робить його систематизацію і класифікацію головоломкою для вченого: розпливчастість меж між його системами, дискретність яких не викликає сумнівів, якщо брати до уваги тільки типові випадки. Навіть між чотирма формами первинного ряду систем, як не капітальні їх відмінності, є переходи. Скажімо, у заростях очерету, пирію, мятлика лугового неможливо відмежувати, «індивіди», особини тут плавно «переходять» в колонію, популяцію і фітоценози. Бамбук-сіянець може розростатися подібним чином навіть на десятки гектарів…
За мільярди років кожна з цих форм зазнала величезних еволюційних перетворень. Ускладнювалися структурні компоненти в організмів. Спочатку нуклеотиди і макромолекули — потім-органели, вже на одноклітинному ступені досягли великої різноманітності. Потім у одноклітинних урізноманітнюються форми життєвого циклу. У багатоклітинних поступово виникає складна система тканин, органів і їх блоків. На новій основі розчленовуються фази життєвого циклу. І так далі.
Великі етапи еволюції слід виділяти і при розгляді популяційно-видової, біоценотичної та біосферної форм існування живого. Але, на жаль, добре вивчені нині лише головні ступені еволюції організмів, багато гірше справа йде з видовою формою, а системи ж біоценозів і біосфери вивчені і зовсім погано.
З усіх цих форм існування життя особливо цікава для еволюціоніста популяційно-видова форма. Справа в тому, що тільки ця форма може бути полем діяльності природного відбору. Макромолекула, клітина, орган, вищий організм — жорстко детерміновані системи. Їх блоки настільки спаяні, що всередині них природний відбір діяти не може. Інша справа — компоненти популяції: з одного боку, вони пов’язані «не намертво», з іншого — досить залежні один від одного.
Склалося так, що саме популяція і вид стали специфічною системою зв’язків між молекулярними, клітинними, тканинними, організмовими, сімейними та іншими структурами.
Важливо ще, що здатність виду до еволюції — лише можливість. Можливість, що не гарантує її реалізації. Подальше залежить від характеру взаємодій виду з середовищем, при цьому головне тут — внутрішня організація самого виду. Ще Дарвін звернув увагу на те, що широко розселені і політипічні види, довівши свої достоїнства в минулому, мають більше шансів процвітати і в майбутньому.
Оскільки вид може перебувати невизначений час у стійкому стані і в той же час еволюціонувати, це поняття внутрішньо суперечливо. Вид — це результат еволюції, і тут він виступає у відносно стабільному стані, якісно певний, цілісний, пристосований до середовища, стійкий, відокремлений від інших груп; але вид — це і вузловий пункт й активний носій еволюції, і тут він менш певний, лабільний, володіє розпливчастими межами.
Слід зазначити, що утворення, яке досягло популяційно-видового рівня, має набагато більш розпливчасті обриси і набагато більш різноманітне за проявами, ніж ідеал виду, за визначенням систематиків. Лише деякі з мільйонів популяцій і рас, що пройшли випробування відбором, досягають рівня “гарного вигляду” , лише деякі виявляються довгоіснуючими, ще менше число видів здатне до тривалої еволюції і до утворення “пучків” нових груп.
Суперечність, що лежить в основі виду, тимчасова і просторова,— основа помилок і однобічних тлумачень цього явища дослідниками. В цілому зараз вже ясно, що без вивчення статики виду неможливо зрозуміти його еволюцію, що вид це необхідна форма, в якій здійснюється життя. Це не означає, правда, що суперечки навколо проблеми виду заглохли, як, втім, навколо всіх інших проблем еволюційної теорії.
Рушійні сили
Отже, арена діяльності еволюційних сил — популяція і вид. Самі ж рушійні сили еволюції – результат взаємодії дуже багатьох факторів. З них, однак, необхідних і загальних, таких, без яких не може йти еволюційний процес, небагато. Відкриті вперше Дарвіном основні фактори нині добре вивчені і підтверджені. Матеріалом для еволюції служить спадковість і мінливість. Єдиним спрямовуючим її фактором — природний відбір як результат боротьби організму за існування. У свою чергу результат відбору – пристосованість його до середовища. Проблему пристосування до середовища — проблему адаптації можна вважати центральною проблемою еволюційного процесу.
Виглядає ця проблема так: не всяка мінливість утримується в поколіннях, а лише та, яка найбільш доцільна в даних умовах. Так виникла пристосованість організму до середовища, організмів один до одного і частин організму — до цілого. На цю особливість живого звернув увагу ще Ламарк. Але він вирішив просто – організмам спочатку властива здатність змінюватися доцільно, прагнення до прогресу “заздалегідь” задано. Таким чином, проблема була знята. Її замінило чисто мольерівське пояснення – масляність властивість масла, – яке, на жаль, допускають вчені і зараз.
Чарльзу Дарвіну вдалося приступити до вирішення цієї проблеми. Засновник еволюційної теорії — на відміну від Кюв’є і Ламарка — прийшов до висновку, що пристосованість організму відносна, а не абсолютна, і довів, що ця властивість не може не бути відносною, так як механізм еволюції пов’язаний з відбором невизначених, ненаправлених змін; як тільки відбір перестає вдосконалювати організм, не змінюється більше і ступінь його доцільності.
Правда, багато послідовників Дарвіна і біологи більш пізнього часу на цій підставі дійшли помилкового висновку, що ступінь пристосованості зовсім не розвивається в часі, що у вищих і нижчих, у бактерій і людини, вона однакова. Виживають адже і ті, і інші.
І ось якось англійцю Ту Дею прийшов в голову дуже простий спосіб кількісного вимірювання ступеня доцільності: по виживаності, тобто відношенням числа народжених до числа померлих. У прогресивному ряді живих істот число народжених падає. Але падає надзвичайно і смертність – виходить, що “внесок” кожної особини в прогрес популяції збільшується. І ось з’ясувалося, що «економічність» живого протягом мільярдів років еволюції виростає на одинадцять — чотирнадцять порядків величин. Іншими словами, на чотирнадцять порядків менше ймовірність того, що одна середня особина виду бактерій внесе вклад в життя свого виду, ніж одна особина мавпи, антилопи та інших…
Однак повернімося до логіки міркувань Дарвіна. Наступним його висновком було: кількість народжених у будь-якого виду завжди перевищує кількість доживших до статевої зрілості. Сукупність причин, що викликають це падіння чисельності, він називав боротьбою за існування.
Тепер ця метафора в результаті робіт екологів, біоценологів і експериментальних еволюціоністів отримала розшифровку. Ми добре знаємо і умови конкурентної боротьби і стимулюючої або, навпаки, «вбиваючої» дії маси різних зовнішніх для організму і внутрішніх умов його існування. Народженому поколінню протистоять хижаки, паразити, конкуренція, погода, клімат, землетруси і так далі. В результаті залишають потомство лише особини, що пройшли природний відбір.
У свою чергу, як припустив Дарвін далі, відбір буває підтримує або перетворює. Тобто загибель або процвітання частини виду може носити вибірковий характер: може вижити «середня» характерна для виду особина, або, навпаки, «крайні» варіанти. Залежно від цього вид або стабілізується, або змінюється, розвивається в якомусь одному напрямку або декількох відразу, якщо він представлений багатьма популяціями.
Такі, в самому короткому викладі, «основи дарвінізму» — вчення про фактори еволюції. Щоб уявити собі, як йде в цій області справа зараз, потрібно згадати про основні події в біологічній науці, що відбулися за століття, що відокремлює нас від Дарвіна.
Як відомо, спадковість і мінливість в епоху Дарвіна були загальними поняттями. Механізм спадкової мінливості був відкритий завдяки успіхам нової науки — генетики, яка виникла спеціально як галузь знання, що його вивчає. Що це за механізм, тепер всім відомо — це мутаційний процес, закони якого добре вивчені, плюс рекомбінації на всіх рівнях.
Відкриття генетики були, як тепер ясно, абсолютно необхідні для розвитку еволюційної теорії, і зараз здається дивним факт, що швидкий прогрес цієї науки послужив поштовхом до опозиції її дарвінізму.
Причиною зіткнення менделевської генетики з дарвінізмом, як відомо, стали висновки Ст. Іоганнсена з серією дослідів з чистими і змішаними лініями квасолі, які нібито довели, що природний відбір не може грати творчу роль, створити що-небудь нове. Іншим поштовхом до пожвавлення антидарвінізму виявилися дані Де Фріза. Цей дослідник спостерігав у рослини енотери так звані макромутації, що ведуть до великих змін в організмі.
Захопившись відкриттями, які йшли тоді одне за іншим, генетики перестали надавати вирішальне значення природному відбору, вирішили, що хід історії живого переконливо пояснює одна лише спадкова мінливість. В результаті з’явилася ціла група недарвіністських теорій на кшталт мутаціонізму і т. п.
Генетики, критикуючи класичний дарвінізм за поступки ламаркізму, були переконані, що ламаркістські тенденції притаманні йому органічно. Як не дивно, ні послідовні дарвіністи, ні їх противники не помічали, що при «прямому» успадкуванні набутих властивостей природний відбір теж виявляється «непотрібним» природі.
З іншого боку, продовжував розвиватися цілий комплекс традиційних наук, що накопичують матеріал про різні системи організму і їх функції. Фізіологія, ембріологія і інші галузі, розвиваючись у відриві від генетики і розглядаючи унікальну пристосованість організмів до середовища «зі своєї дзвіниці», стали звертати увагу тільки на пристосувальну роль, адаптивність виникаючих змін. Звідси виникло переконання, що еволюцію можна пояснити прямим впливом середовища на організм і успадкуванням набутих ознак,— неоламаркізм.
Паралельно час від часу оживали в біології і телеологічні, віталістські концепції, що беруть початок ще від Аристотеля, але в наш час знаходять собі грунт в областях, де накопичувалися факти, не отримуючи пояснення. А таких областей в біології ставало все більше. Перед дослідником все в нових і нових ракурсах відкривався чудово налагоджений, найтоншим чином відрегульований живий світ, а в поясненнях його походження було ще досить багато провалів, і, що гірше, ледь закривалися одні, з’являлися нові.
Довгий час в після дарвінівський період концепції природного відбору не вистачало фактів. Вона змушена була спиратися на аналогії з штучним відбором, на непрямі докази, пов’язані з дослідженнями не самого природного відбору, а його результатів.
Але поступово почали ставитися експерименти з вивчення факторів еволюції, даних накопичувалося все більше, головні ж таки дані стала постачати сама генетика. З цього почався новий синтез еволюційної теорії. «Еволюція, сучасний синтез» так називається книга Дж. Хакслі, після появи якої багато хто став називати нинішній етап у розвитку дарвінізму синтетичною теорією еволюції. Книжка вийшла під час Другої світової війни, початок ж новому етапу було покладено багато раніше — роботами популяційних генетиків, з одного боку, біогеоценологів — з іншого, і фізіологів та ембріологів — з третього.
Етап цей характерний вивченням мікроеволюційних процесів, що йдуть всередині популяції, пускових моментів еволюції, початкових стадій пристосування і формоутворення, які, врешті-решт, ведуть до видоутворення.
Займається синтетична теорія і проблемами макроеволюції, вважаючи, що вони принципово не відрізняються від тих, що відносяться до подій всередині виду, і розглядаючи ці два процеси як дві сторони єдиного процесу еволюції.
Переважна більшість біологів, так чи інакше, підтримує зараз цю теорію, тобто сучасний дарвінізм.
Дарвінівські фактори еволюції в “перекладі” на нинішню біологічну мову виглядають так: мутації, ізоляція, популяційні хвилі, природний відбір. Щоправда, одностайно зізнаються лише два з них — два “середні” вже предмет дискусії. Ізоляція — явище складне, є багато її видів: географічна, екологічна, фізіологічна, поведінкова.
Популяції, що живуть на різних островах, ізольовані географічно, популяції — що мешкають на одному лужку, – але час цвітіння у них різний – розділені еколого-фізіологічно.
Навколо значення фактора ізоляції для видоутворення йдуть суперечки. Скажімо, Ернст Майр, один з стовпів синтетичної теорії, насилу визнає можливість видоутворення без географо-екологічної ізоляції. А між тим видоутворення організмів, що живуть пліч-о-пліч, може благополучно йти, якщо в популяції утворюються феногрупи з різними, наприклад, термінами розмноження.
Такий фактор, як популяційні хвилі, визнається теж не всіма, хоча вважається найважливішим багатьма корифеями синтетичної теорії: екологи встановили, що чисельність популяцій коливається, в одні роки багато хутра, інші — яблук, треті, скажімо, неймовірна кількість лободи; і час цих коливань буває різним — від місяців до століть, різна у різних видів і величина коливань.
Так от зміни чисельності змінюють концентрацію мутацій в популяції, а це важливо для подальшого видоутворення. Суперечка йде лише про те, чи є популяційні хвилі самостійним фактором еволюції або наслідком інших.
Серед “загальновизнаних” факторів, треба сказати, зустрічаються дуже важливі. Одна з особливостей синтетичної теорії еволюції — якраз та, що вона «привласнила» (бо вона, до речі, так і називається) деякі з факторів, які фігурували раніше навіть в антидарвінівських теоріях. Правда, зараз вони постають не зовсім в тому вигляді, в якому виглядали «в руках» антидарвіністів.
Так, американський біолог Ч. Девенспорт і француз Л. Кено півстоліття тому висунули концепцію так званого преадаптаціонізму, він спирається на вже згадувані досліди Де Фріза, що виявив макромутації.
Як всі антидарвіністи, преадаптаціоністи відкидали природний відбір, припускаючи процес еволюції в такому вигляді: у крота, скажімо, спочатку виникає відсутність очей і риюча конструкція кінцівок, а потім звір ходить по світу, шукаючи підходяще середовище проживання і спосіб вживання нового органу.
Ця теорія в цілому була неспроможна, але вчені довели сам факт існування мутацій, що виникли як готове пристосування. І преадаптація стала одним з факторів еволюції в синтетичній теорії. Фактор цей діє, якщо пристосування виявляється згодним із середовищем, і не діє, якщо умови несприятливі. Воно не є самодостатнім і не суперечить природному відбору. Але не враховуючи ролі преадаптації як певної фази в еволюційному процесі, ми не можемо пояснити появи жодного великого нововведення, так як з’ясувалося, що саме великі нововведення мають преадаптивну фазу.
Скажімо, в дельті річки падає градієнт солоності; якщо розглянути які живуть в цих місцях популяції риб, крабів або раків, то з’ясується, що деякі з морських тварин поступово набувають здатність переносити прісну воду і, навпаки, деякі прісноводні, запливаючи в море, знаходять можливість жити в солоній воді. Так ось, виявилося, що в популяціях, скажімо, прісноводного виду існують мутанти, які заздалегідь здатні переносити більшу солоність, ніж мільйон їхніх побратимів. В умовах прісної води ця мутація нейтральна, вона “чекає” своєї години і при змінах умов знаходить вирішальну перевагу.
Шляхи, по яким слідує життя
Залишилося розглянути, що ж виходить в результаті дії головних еволюційних факторів. Які шляхи вибирає собі життя?
Одна з головних встановлених наукою закономірностей — адаптивний характер процесу еволюції. За значенням дорівнює їй незворотність цього процесу.
Як і все в нашому світі, закон цей не має абсолютного значення. Так, теоретично допустимо, що у монозиготної лінії може виникнути одна генна мутація, а потім — мутація зворотна по цьому самому гену, тоді перед нами — чиста оборотність. Але ж таке практично ніколи не станеться, бо треба ще, щоб одночасно весь гігантський організм перебував при цьому в стані досконалого спокою, ніяк не змінювався.
Третя закономірність — нерівномірність темпів еволюції. Бувають періоди, коли умови середовища для даного виду не змінюються, група стабілізується, і природний відбір, як вже зазначалося, відкидає всі крайні «варіанти», підтримує «середню» особину, тип виду. Тоді — часом протягом мільйонів років — буде підтримуватися статус кво.
Процес, який ще Томас Гекслі назвав персистуванням, – «підвид» закону нерівномірності розвитку, і стосується він як цілих груп, так і окремих ознак організму. Скажімо, енергетична основа будь-якої клітини — АТФ і деякі інші речовини практично не змінюється, можливо, вже мільярд років. Те ж можна сказати про ознаки, що відносяться до зародків вищих організмів. Адже якщо якась деталь ембріона зміниться, то це зазвичай так розрегулює – за принципом підсилювача – дорослий стан особини, що їй загрожує чи не стовідсотковий смертельний результат. Волею-неволею через смертність всіх дорослих форм, що носять ембріональні зміни, такі ознаки виявляються досить стабільними. І темп еволюції ембріонів в сотні разів нижче, ніж темп еволюції дорослих форм.
Квантова еволюція
Поки ми говорили про випадки, коли еволюція завмирає. Але знаємо ми і прямо протилежні факти — так званої квантової еволюції. Термін цей ввів американський еволюціоніст Сімпсон.
Типовий приклад великих разових перетворень — поліплоїдія на основі віддалених схрещувань, гібридизація у рослин. Це міжвидові, міжродові схрещування, при яких часто виникає такий геном, який складається з геномів батьків і далі не розщеплюється. Виходить тетраплоїд, октаплоїд і так далі, а фенотипічно це новоутворення, яке поєднує ознаки різних родів і свої «власні» нові ознаки. В останні п’ятнадцять років вивчені гібрідогенні цикли у рослин, коли особливо агресивний генотип при схрещуванні поглинає навіть кілька геномів і об’єднує разом п’ять-шість не те що видів, а родів. Це відбувається порівняно швидко, за десяток років, тобто за життя одного покоління спостерігачів. Нова форма при цьому вже заздалегідь пристосована до існування, зрозуміло чому — адже вона «складена» із видів з ознаками, вже відібраними природою.
Темп еволюційного процесу в таких випадках надзвичайно прискорюється. Природно, квантова еволюція не суперечить дарвінізму, хоча спочатку здавалося, що відбору в цьому випадку немає місця…
Інший клас закономірностей еволюції – напрямки або основні її шляхи. Проблему цю Дарвін ще не міг поставити в загальному вигляді, і тому вона довгий час була буквально заповідником всяких метафізичних теорій, поки основи вчення про головні напрями еволюційного процесу не розробив Олексій Миколайович Северцов.
Северцов вивчив морфо-фізіологічні шляхи, за допомогою яких популяцією може досягатися стан процвітання. Він виходив з положення, що вся еволюція адаптивна. Придбання популяцією всякого нового пристосування призводить до того, що вона починає збільшувати чисельність, розселяється за межі свого екологічного та географічного ареалу і, нарешті, розпадається на підгрупи. Отже, множення, розповзання і розпад. Все це було об’єднано під назвою «біологічний прогрес». Якими ж морфологічними шляхами досягається такий стан?
Їх три.
- Перший — ускладнення організації, інтенсифікація обміну. У цьому випадку біологічному прогресу сприяє морфо-фізіологічний, який Северцов назвав “ароморфоз“. Нині термін цей уточнений, і процес названий арогенезом.
- Другий шлях – його знав ще Мечников – це алогенез, перепристосування, шлях без підвищення або зниження рівня організації. Приклад такого шляху еволюції — безліч видів антилоп в Африці, рівних за своїм рівнем і пристосованих кожен до якихось конкретних умов — савани, степу, пустелі і т. д.
- Третій шлях — коли біологічний прогрес досягається шляхом морфо-фізіологічного регресу. Багато вважають, що віруси, скажімо, це не нащадки найдавніших живих організмів, а спрощена група, що отримала в результаті гігантські переваги.
Якщо поглянути на ці шляхи з екологічної точки зору, легко переконатися, що арогенез збільшує широту пристосованості: скажімо, холоднокровна стало теплокровною твариною — широта проживання її стала більшою, незалежність від середовища збільшилася; інстинктивна поведінка змінюється розумною — можливості розширюються. Алогенез лише змінює середовище; Луки – лісами або степом: рівне на рівне. Третій же шлях в екологічному ракурсі – це спеціалізація, а не регрес. Вона означає зазвичай зменшення числа функцій з інтенсифікацією залишених. Наша кінцівка, скажімо, п’ятипала, примітивна з примітивних. В ній збереглася стегоцефальна ознака, ознака першої з сухопутних тварин. Копито ж у порівнянні з нею зазнало більше змін, але воно розвивалося в бік спеціалізації, ряд функцій у цього органу відпав, одна ж інтенсифікувалася.
Ще один клас закономірностей деякі дослідники теж називають напрямками еволюції. (Взагалі з термінологією в цій справі поки ще немає визначеності. Щоб краще уявити, про що йде мова, знову-таки повернімося до витоків еволюційного вчення. Ламарк, створюючи першу в історії біології еволюційну теорію, уявляв собі еволюцію як процес, який задовольняє принципу суворого паралелізму. З найдавніших часів і донині – по Ламарку – живе з’являється від неживого, довільно самозароджується життя. Виниклі істоти під впливом закладеного в них природою прагнення до прогресу розвиваються зі щабля на щабель, поки не досягнуть вершини. Вершина ця – людина. Така схема, телеологічна по суті, якщо її зображати графічно, нагадує житнє поле: все, що з’явилося, йде вгору.
Дарвін, спостерігаючи за діями селекціонерів, встановив, що вони створюють багато порід тварин і сорти рослин з небагатьох основних форм, основа селекції — рух від одноманітності до різноманітності. Цей принцип, названий ним дивергенцією, Дарвін переніс на всю природу.
Кілька десятиліть тому це первісне формулювання була дещо перейменоване і розширене, стали говорити про адаптивну радіацію. Але суть цього твердження в даний час повністю зберігається. Воно було підтверджено експериментами і спостереженнями на культурних і на диких видах. Принцип дивергенції лежить в основі спостережуваної нині різноманітності. Принципи відбору, на підставі яких відбувається процес дивергенції, відомі. Але є і зараз, особливо серед палеонтологів, прихильники спочатку паралельного і навіть конвергентного (від різного до подібного) розвитку живого. І хоча в ортодоксальному вигляді ця позиція спростовується численними фактами, що ілюструють дивергентне походження сучасного розмаїття видів, вести боротьбу з нею слід все ж без особливого запалу.
Дуже мало ми знаємо про перед біологічну еволюцію, і дуже можливо, що поява життя була не строго монофілетичною, що воно виникло в різних точках або в декількох варіантах відразу або що цей процес починався кілька разів.
Те ж саме можна сказати і про прояви паралелізму і конвергенції – сходження в біографії будь-яких великих окремих груп живого. Так, Татарінов, працюючи над звіроподібними ящерами (теріодонтами), знайшов дуже цікаві факти, які підтвердили, що паралелізм і конвергенція грають насправді більш значну роль в еволюції, ніж колись уявляли собі це класики дарвіністської палеонтології.
Як витягти закон з «викопного» стану!
Нарешті, поставимо собі таке питання: а яке джерело інформації про еволюцію? Адже прямому спостереженню еволюційний процес недоступний. Картина еволюції складається на підставі залишків організмів і слідів їх життєдіяльності, що перейшли у викопний стан. Але ж жоден фактор еволюції у викопний стан не переходить… Це і створює колосальну кількість протиріч в біології і перш за все труднощі з методами.
Неможливо “підглянути”, як йде еволюція; процес її розтягнутий в часі і прихований від очей. Досліднику доводиться монтувати «фільм» про еволюцію з нескінченної безлічі нерухомих кадрів — спостережень. При цьому ще не відомо чи не завжди відомо, які з них дублюють один одного, куди який кадр ставити, які з них взагалі «не звідси», які зняті невірно або не в тому світлі, і часом виникає сумнів, чи з початку показується фільм. Не вдаються поки і «мультиплікації» — моделі процесу. Занадто він складний для них і занадто багато в ньому невідомих.
Вже давно прихильники самих різних напрямів еволюційної теорії зійшлися на тому, що єдиний спосіб дослідження — вивчати закони, нині чинні, і обережно екстраполювати їх в минуле. Це метод актуалізму.
Чинні нині фактори та їх взаємодію наука досліджує експериментально. Експерименти ці відтворювані. Словом, наукова робота ведеться звичайним для сучасного природознавства способом. В результаті склалося повністю обгрунтоване вчення про мікроеволюції, тобто про еволюційний процес, що йде всередині видів. Не можна сказати, що вчення це вже завершено, перед вченими ще стоїть безліч труднощів, але комплекс точних уявлень в цій області вже є. Можна навіть стверджувати, що нині чинні чинники еволюції досліджені.
Труднощі полягають в екстраполяції. У проблемі, як далеко ми можемо «заходити» в минуле, і чи маємо ми взагалі на це право. Дарвін мовчки, а потім, коли він ближче познайомився з геологічними даними, вже і «вголос» стверджував вічність законів природи; той, що діє нині, на його думку, діяв і в минулому.
Така точка зору викликала великі заперечення вже в XIX столітті. Серед геологів, палеонтологів і біологів-неокатастрофістів, послідовників Кюв’є, знайшлися її противники. Було висловлено багато «зайвого», але одна з ідей опонентів виявилася в цілому правильною — думка про те, що самі еволюційні закони теж еволюціонують, зазнають зміни.
Легко припустити, що взаємодія факторів еволюції три мільярди років тому була зовсім іншою, ніж зараз, і у бактерій зараз вона інша, ніж у мавп, у рослин — інша, ніж у тварин. Своєрідність організації визначає і своєрідність розвитку. Адже якщо, скажімо, у організму спадкова речовина ще навіть не організована в хромосоми, то й фактори еволюції такого організму будуть проявлятися по-іншому, ніж у вищих тварин. Наприклад, відбір виявиться не настільки точним, інша буде мінливість і так далі.
І тому — лише якщо ми говоримо про порівняно недавні часи та про тих таксонів, які представлені і в наш час досить повно, всі сто відсотків біологів проголосують за дозволеність екстраполяції. Але якщо йдеться про мільярд років, тобто коли ми переходимо до палеозою або протерозою, до тих часів, до яких належить виникнення великих груп — типів організації живого, нас можуть чекати несподіванки.
На цій підставі була висловлена не раз точка зору, що дарвінізм правильний тільки для сучасності, але зовсім непридатний для далеких епох.
В цілому такий висновок був би невірний. Як показали результати численних досліджень, проведених на різних сучасних групах тварин і рослин, яких ми з повним правом можемо розглядати як представників минулих історичних періодів, еволюційний процес базується на природному відборі. Однак сам природний відбір під впливом інших факторів, що змінюються з часом, виступає в різних варіантах.
Ми не можемо ще з достовірністю встановити повну картину еволюції, ми лише намацуємо якісь її основи, тому ми повинні з особливою дбайливістю ставитися до всіх фактів, до всіх деталей, якщо тільки це насправді деталі реальності, а не гіпертрофоване, спотворене їх тлумачення.
Як не дивно, відокремити факти від їх передчасного пояснення стало істотним завданням еволюціоніста. Хотілося б, щоб за всім сказаним проглядалася думка про те, що еволюційна теорія – незакінчена система, що це сукупність не істин в кінцевій інстанції, а проблем, що знаходяться в роботі.
Природно, не всі з цих проблем «освоєні» теорією в рівній мірі. Еволюційні біологічні науки — «постачають» еволюційній теорії дані, фактичний матеріал, розвиваються нерівномірно, як, втім, взагалі вся наука. Тому теоретичним посиланням часом гостро не вистачає фактів, і, навпаки, деякі нововиявлені факти теорія буває не в змозі відразу пояснити, і виникають різні точки зору. Вже наводилися подібні приклади з історії еволюційного вчення. У наші дні переконливу картину “хвороби росту” представляє собою, наприклад, проблема внутрішніх факторів еволюції, їх ролі в її процесі.
«Головне заперечення, яке висувається проти синтетичної теорії еволюції, – пише Ернст Майр, – полягає в її нездатності нібито пояснити деякі еволюційні явища, перш за все спрямовану еволюцію, паралелізм і еволюційні тенденції». Звичайно, проблема внутрішніх факторів еволюції не може бути зведена до її спрямованості, оскільки сама спрямованість — результат взаємодії багатьох факторів, в числі яких певну роль відіграють і зовнішні, підсумок суперечливих відносин живого з навколишнім середовищем. І для розуміння спрямованості треба враховувати еволюційне минуле кожної групи організмів, шлях, по якому вони розвивались. Саме він визначає подальший еволюційний шлях, накладає обмеження. Грубий приклад: скажімо, у мавпи ніяким чином не може виникнути такої мутації, в результаті якої у неї виросли б пір’я.
В кожному організмі є ціла ієрархія «заборон» — від генетичних, фізіологічних, онтогенетичних до поведінкових. І всі вони у всіх своїх складних взаємовпливах, в кінцевому рахунку, зумовлюють шлях, по якому стане розвиватися він далі. Але приречення це не жорстке, воно «залишає місце» для варіантів, мало того, саме «передбачає» їх обов’язковість. Так, апарат, що регулює в організмі мутаційний процес, під впливом обставин, іноді і зовнішніх, часом перебудовується на інший режим роботи», і разом з ним змінюють характер і самі мутації.
Розмірковуючи про цю проблему, не можна не віддати належне чудовій ідеї Івана Івановича Шмальгаузена про стабілізуючий відбір. Це, мабуть, найбільше доповнення до еволюційної теорії після вчення про популяцію. Іван Іванович розглядав еволюцію саме як регульований процес і дав наукове обґрунтування ідеї про закономірний його характер. Він парадоксальним чином поставив в центр міркувань не змінюваність органічних форм, а їх стійкість, здатність в мінливих ситуаціях зберігати “норму”. Ось цей “динамічний режим” , сам по собі підсумок еволюції, на його думку, направляє її подальший перебіг. Таким чином, Шмальгаузену вдалося подолати протиріччя, на яке наштовхується еволюційна теорія: адже мутація «позитивна», щоб не виявитися смертельною для організму, повинна змінювати його гармонійно. І це дійсно буває так, скажімо, якщо у птиці змінюються крила, то відразу обидва, захисне забарвлення з’являється на спині і т. д.,— але вимагає спеціального пояснення, і Шмальгаузен таке пояснення дав: гармонія ця — результат виживання тих особин, які розвиваються на більш досконалій системі внутрішніх регуляцій.
Ідея стабілізуючого відбору має велике майбутнє, так як на внутрішні фактори еволюційного процесу стали звертати спеціальну увагу переважно останнім часом. Раніше ж багато дарвіністів “класичного” періоду недооцінювали їх, всю увагу приділяючи «боротьбі за існування», тобто зовнішнім чинникам розвитку.
Але Іван Іванович Шмальгаузен, природно, лише поклав початок вирішенню проблеми внутрішніх факторів і спрямованості еволюції, а тому вона досі викликає найбільші розбіжності і різнотлумачення, іноді і в телеологічному дусі.
Мабуть, немає сенсу перераховувати інші теми еволюційної теорії, що викликають суперечки. Але варто повторити: різні точки зору виникають або при сильній диференціації знань без їх інтеграції, і тоді кожен починає судити «зі своєї дзвіниці», або коли фактів не вистачає або їх важко добувати, або, навпаки, коли їх занадто багато, а не вистачає теоретичних обґрунтувань, або, нарешті, під впливом впровадження в біологію небіологічних наук зі своїми методами. Про це, до речі, досі не говорилося, і варто навести коротенький приклад.
Останнім часом часто робляться спроби оцінити еволюційне вчення з позиції таких наук, як кібернетика та біофізика. Так, Ст. Бір, кібернетик, який займається системами управління виробництвом, вважає, що на основі випадковостей еволюція «протікала б надто повільно і дуже ненадійно, щоб з їх допомогою можна було пояснити, як відбувається будь-яка адаптація». В управлінні органічної еволюції він бачить ті ж системи передетермінованості і планування, що і на виробництві. Треба сказати, що інші його колеги — Н. Вінер, Р. Ешбі – цілком підтримують дарвіністську “програму” зі своїх кібернетичних позицій. Відомі математичні моделі еволюції, побудовані Ешбі, які підтверджують, що її рушійна сила — природний відбір.
На закінчення хочеться сказати, що з усього різноманіття еволюційних вчень лише теорія природного відбору зберігає своє наукове значення вже більше століття, продовжує розроблятися далі і в даний час більшістю дослідників визнана основою дослідження причинних механізмів еволюції. Але її розвиток на всьому протязі історії дарвінізму супроводжувався дискусіями і виступами опонентів, які різко загострювали постановку ще не вирішених питань. Найчастіше подібні загострення йшли «на користь справі», до того ж деякі з висновків опонентів потім ставали надбанням теорії.
Автор: Б. С. Соколов.